FARMACÉUTICOS N.º 392 -
Febrero
2014
42
Asesoramiento Farmacéutico
– Prostaglandinas (PG) que, pese a carecer de efecto directo,
potencian los efectos algógenos de la serotonina y la bradicinina
– Cininas: péptidos producidos por escisión proteolítica a partir
de precursores plasmáticos inactivos: bradicinina
– Sustancias exógenas: capsaicina
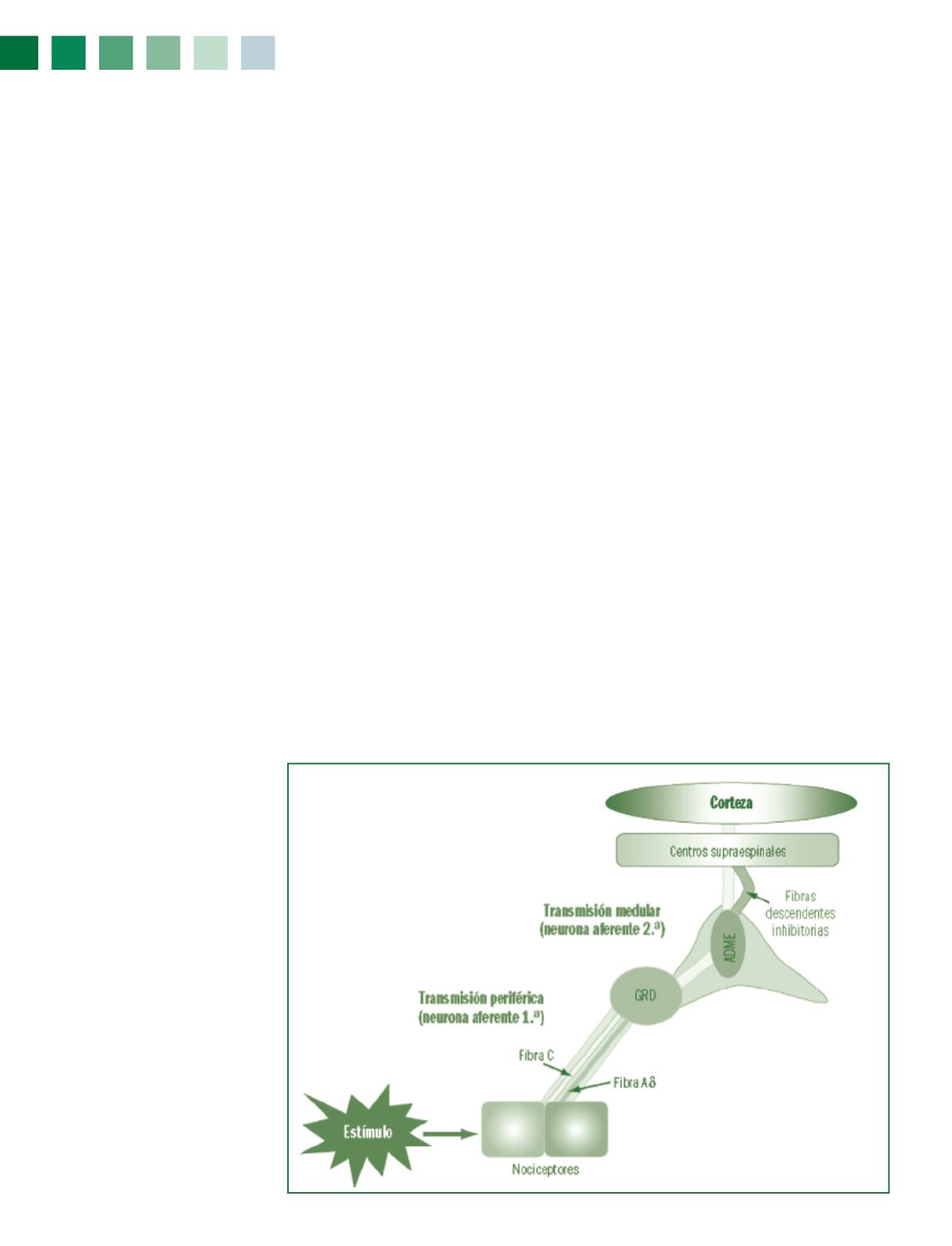
En respuesta a la estimulación, los nociceptores producen y
liberan mediadores químicos de acción rápida. Generalmen-
te, se trata de aminoácidos (ácido glutámico) o pequeños pép-
tidos (cadenas de hasta 25 aminoácidos), tales como la
sus-
tancia P
, formada por 11 aminoácidos y considerada como el
principal neurotransmisor nociceptivo en las fibras de tipo C.
Estos mediadores químicos inducen modificaciones del flujo
iónico (fundamentalmente, salida de potasio y entrada de cal-
cio) que conducen a la despolarización de la membrana neu-
ronal. Si el nivel de despolarización es adecuado, se genera-
rá un potencial de acción, que se propaga en sentido aferente
(desde la periferia hacia las estructuras nerviosas superiores).
El impulso nociceptivo recorre los nervios periféricos, llega a
las capas superficiales del asta dorsal espinal y asciende por
alguna de las múltiples vías medulares, pasando por el cere-
bro medio, para acabar en el tálamo, desde donde se distribuye
hacia la corteza cerebral.
El estímulo nociceptivo activa tanto los sistemas ascendentes de
transmisión del dolor como los sistemas endógenos inhibitorios
de la transmisión nociceptiva (opiáceo,
α
2
-adrenérgico, colinér-
gico, etc.), situados a nivel periférico, espinal y supraespinal. La
integración de la transmisión
excitatoria e inhibitoria a estos
tres niveles determina las prin-
cipales características de la
transmisión y percepción del
dolor y permite al sistema ner-
vioso discriminar entre estí-
mulos lesivos y otros inocuos.
El impulso nociceptivo
es transmitido por diversos
tipos de conducciones nervio-
sas (formadas, obviamente,
por neuronas). La velocidad
de transmisión del impulso
varía según el grado de mie-
linización de estas conduccio-
nes, al actuar la mielina como
un auténtico aislante eléctri-
co. Así, se ha estimado en un
valor de 6 metros por segundo
(6 m/s) por cada micra (µm) de grosor de la capa de mielina.
Dado que el diámetro neuronal es de 2 a 20 µm, la velocidad
de conducción fisiológica de los estímulos dolorosos está com-
prendida entre 12 y 120 m/s. En casos especiales, el diámetro
neuronal alcanza valores tan bajos como 0,2 µm, lo que implica
velocidades de transmisión de apenas 0,8 m/s. Este tipo espe-
cial de neuronas juega un papel decisivo en la transmisión de
los impulsos que dan origen al
dolor lento
, y son denomina-
das fibras C.
Las fibras con mayor grosor en la capa de mielina (entre 1 y 6
µm) son denominadas fibras A
δ
(léase A “delta”), que conducen
los impulsos causantes del dolor rápido (la velocidad de trans-
misión es de 6 a 35 m/s). Este último tipo de fibras son las úni-
cas activadas en los procesos en los que hay un intenso estímu-
lo cutáneo como, por ejemplo, el que se produce por el pinchazo
de un alfiler.
Según el tipo de fibra, el acceso al tálamo se produce por vías
medulares diferentes. El complejo ventrobasal del tálamo (forma-
do por los núcleos laterales y posteriores) recibe conexiones neu-
ronales procedentes del sistema de conducción rápida. En estos
núcleos están representados topográficamente la cara, la cabeza
y el cuerpo, lo que implica un elevado grado de especialización y
selectividad en la ulterior interpretación de los impulsos doloro-
sos a nivel de la corteza cerebral.
Las neuronas que siguen el sistema ascendente múltiple de la
médula transmiten los impulsos lentos (dolor lento), pasan por


